15. A Bit Like a Line Dance (945)
- lscole
- Apr 30, 2025
- 4 min read
Updated: Jun 17
Having just reviewed all the steps in the cell cycle, we now focus on the fourth step, mitosis, where the duplicated genome is separated into two new cells. I liken this amazing feat of molecular choreography to a Western-style line dance.
Let's review mitosis from start to finish before covering each step in more detail.
Our starting point will be the completion of interphase--that is, phases G1, S, and G2. The genome was copied in S phase. All the chromosomes in the nucleus are now in the form of long, stringy DNA.
In the first phase of mitosis, prophase, the stringy chromosomal DNA condenses into compact, visible structures.
They then make their way to an imaginary line at the center of the cell known as the equator.
There, molecular cables attach each to structures at the two poles of the cell (metaphase).
The cables then pull the chromosomes toward each pole (anaphase), where the chromosomes de-condense and two new nuclear membranes form around them, creating two new nuclei (telophase).
Finally, the cell pinches in the middle and becomes two cells (cytokinesis).
Let's walk through each of these phases.
Interphase: The Dancers Haven't Arrived Yet
Our starting point is a cell that has passed through G2 after replicating its genome in S phase.
After replication, each of the 46 chromosomes consists of two identical copies joined together. Each of these joined identical copies is called a chromatid.
The chromosomes/chromatids can't yet be distinguished visually. But note an important detail: two small star-like objects float outside the nucleus. These are centrosomes. They'll play a key role in chromosome movement.

Prophase: The Dancers Appear
The stringy chromosomes in the nucleus compact and the star-like centrosomes move to opposite poles. The chromosomes are compacted by protein complexes and then looped and coiled into compact units by other protein complexes.
Next, hundreds to thousands of special ring-shaped proteins are loaded onto the chromosomes and encircle them, holding the two copies (the sister chromatids) together--much like rings around a bunch of wires.
Now the chromosomes are in a form that can be pulled into the new daughter cells. These are our line dancers.
In addition to condensing their chromosomes, cells begin reorganizing their internal scaffolding.
The two centrosomes move to opposite ends of the cell and arrays of microtubules assemble between them. Together, they form the mitotic spindle—a dynamic structure that forms and reorganizes itself. The mitotic spindle will soon capture and move chromosomes.
By the end of prophase, the centrosomes will be positioned at opposite poles with the spindle fibers radiating outward and, in some cases, overlapping across the cell’s center. Two final steps complete prophase.
First, the nuclear membrane breaks down into fragments. These will later be recycled to form new nuclei in the daughter cells. This exposes the chromosomes to the mitotic spindle.
Second, protein hook-like structures called kinetochores assemble at the centromeres. These will act as microtubule docking sites, allowing the mitotic spindle to attach to chromosomes.
What is a chromatid?
Let's pause for a moment to take a look at one of these condensed chromosomes and clarify terminology.

When a chromosome is copied during S phase, the result is two identical DNA molecules that remain attached at a central point called the centromere.
Under the microscope, this structure looks like an “X.” Each half of the X is one of these identical copies. Biologists call each half a chromatid, and together they are referred to as sister chromatids.
Although each chromatid is a complete copy of the chromosome, the pair is still treated as a single unit because the two copies are physically linked. Only when they are pulled apart during mitosis do they become independent chromosomes.
For our purposes, it’s helpful to think of each “X-shaped chromosome” as two identical copies of the same chromosome joined together, waiting to be separated.
Metaphase: The Dancers Line Up
The 46 X-shaped "double chromosomes" line up next to each other at the equator of the cell. The centrosomes--now at opposite ends of the cell--send out additional spindle fibers that attach to the molecular hooks.
By the end of metaphase, all 46 chromosomes (92 chromatids) will be aligned along the equator and each of their hooks will be attached to a spindle fiber emanating from one or the other of the centrosomes.
Anaphase: The Dance Begins
At the start of anaphase, a specialized enzyme within the cell breaks the bonds holding each pair of chromosomes together.
The enzyme responsible waits in an inactive state. When the cell confirms that the chromosomes are properly aligned, the enzyme switches on. It cuts the protein rings linking the chromosomes, allowing them to be pulled apart.
The chromosomes are then pulled toward the centrosomes by a process in which the spindle fibers shorten in a controlled way.
At the same time, other spindle fibers that connect the two centrosomes do the opposite—they lengthen, elongating the cell and preparing it for cleavage.
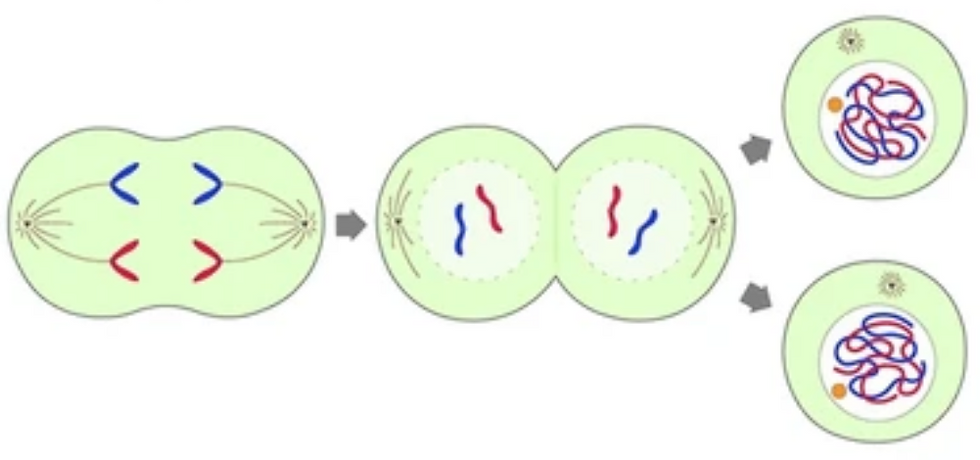
Telophase: Two New Dance Troupes Form
In telophase, first, fragments of the nuclear membrane that were broken down at the end of prophase are recycled to create two new nuclear membranes in the daughter cells.
Second, the cell membrane pinches inward at the center, separating the cell into two daughter cells, each receiving a complete set of chromosomes and the components needed to function.
At the same time, the chromosomes relax back into their extended form within the newly formed nuclei.
What began as a single cell is now two—each carrying an identical copy of the genome.
From this point on, our focus will be the process by which the cell manufactures these two genomes.

Comments